
超普通心理学/感觉与知觉/知觉系统
| 超普通心理学 |
|---|
| 心理学绪论 |
| 心理学研究法 |
| 行为的生理基础 |
| 感觉与知觉 |
| 感觉系统‧知觉系统 |
| 意识 |
| 睡眠‧梦 |
| 学习 |
| 记忆 |
| 感官记忆‧短期记忆‧工作记忆‧长期记忆 |
| 语言与思考 |
| 认知科学‧语言‧思考 |
| 智力 |
| 智力理论‧人工智慧 |
| 发展心理学 |
| 动机与情绪 |
| 情绪的种类 |
| 压力与健康 |
| 性格 |
| 常见的人格障碍类型 |
| 心理异常 |
| 心理异常的治疗 |
| 社会心理学 |
| 社会认知、基模、归因、态度‧自我 |
本章节介绍身体各项知觉。
深度知觉
[编辑]视知觉给我们提供一个重要的能力:即使视网膜成像是二维的,我们仍能看到三维世界。差别在于所接收到的讯息深度不同,分为单眼线索和双眼线索。单眼线索是只需要一只眼睛就能感受的线索;双眼线索是需要同时用两只眼睛才能感受到的线索。若我们将一只眼睛闭起来,我们还是多少能够感知一些深度的讯息,这些讯息被称之为单眼深度线索(monocular depth cues),或者是图画线索(pictorial cues),因为这种线索利用了艺术家在绘制图画时会用到的技巧,如透视线。
部分人患有立体盲,不能利用视网膜的成像差异适当估计距离。最常见的立体盲是斜视,两眼从不同的方向看向物体。这是一种严重的缺陷,可导致叠像的产生,或即使双眼都睁开时,却只靠一只眼睛判断距离。
静态线索
[编辑]- 双眼深度线索(binocular depth cues)
我们的双眼借由两眼视网膜呈像的像差(retinal disparity)以及双眼聚合(convergence)来提供深度讯息。
- 视网膜像差:大脑借由两眼间距所造成其所接收外界影像不同所产生的“像差”,来解读物品在我们这个三元空间中的深度。例如,若有A、B两物体,此两物体在两眼中所见的距离可能因为左右眼视角的不同而有大小的差异,若此AB间的距离在两眼中看起来类似,可能代表AB相距不远;反之,若AB间距离在两眼中有较大的差异,可能代表AB有距离上的不同。
- 双眼聚合(convergence):讲述当我们凝视物体时,双眼瞳孔转向内侧的情况(有点像是轻微斗鸡眼的状况)。若物体离我们十分近,则眼睛需要内转较多才能聚焦在物体上,因此大脑利用来自眼球肌肉的讯息来了解近距离物体的远近。
我们如何把两个不同图像组合或匹配成单一的知觉?按照马尔和葡基耶(Marr & Poggio)的理论,有三条主要原则
- 兼容性原则:为了使两个视网膜成像相配,这两个部分要在某地方是兼容的或相似的,例如颜色和形状。
- 连续性原则:如果两个视网膜像之间的差异与已经建立的邻近视网膜像之间的差异类似,这两个视网膜像更容易匹配。
- 唯一性原则:一个视网膜像的某一个部分仅与另一个视网膜像的部分相配。
- 单眼深度线索(monocular depth cues):
- 质地梯度(texture gradient)
- 距离近的物体将会有较多的细节资讯,其内容是指借由观察递减的纹理来了解距离,简单而言就是因为越往深处,物体质地密度在视觉上会变得越大(这是另一个距离、大小关系所造成的结果),因此可以借由这种“材质纹理”来了解三次元空间中的距离。
- 遮挡(Interposition)
- 对于深度视觉来说,最重要的单眼线索之一是遮挡。当一个近的物体部分的遮住了远处物体时,就出现了遮挡。卡尼撒(Kanizsa)的错觉方块支持了这一个线索存在的想法。它的错觉方块是四颗黑球摆在正方形的四个顶点,然后内侧个被挖去四分之一圆,这个时候,视觉系统在中间产生了一个白色的方块,遮挡在黑球前面,使得圆的缺失部分变的合情合理。
- 直线透视(linear perspective)
- 最简单的例子就是无限延伸的铁轨在视野中的最尽头会似乎会汇合到一个点上,铁轨应该是由两条平行线所组成,可因为庞氏错觉(Ponzo illusion),所以比较接近的物体会看起来较长、而比较远的物体则是看起来较小、较短;
- 熟悉大小(Familiar size)
- 另一个重要的深度线索是熟悉大小。如果我们熟知一个物体的大小,那么,我们就可以用它在视网膜上的大小计算出距离。
- 大气透视法(Atmospheric perspective)
- 大气透视,又称空气透视或空气远近法,是由于受到空气微粒的影响,使得近、中、远距离的物体会因为光学作用而产生不同程度的颜色变化,人们看到近处的景物比远处的景物浓重、色彩饱满、清晰度高。受大气透视影响越严重对比度就会越低,所以也可以说越远的东西失去越多细节表现。正中午的大气透视最弱,清晨及夕阳的斜角光线会让透视感较为显著,而浓雾或严重空气污染会产生更强烈的大气透视效果,例如”伸手不见五指”的情况通常就是浓雾所造成的。这广泛运用在绘画和摄影技巧之中。
动态线索(Motion cues)
[编辑]- 属于单眼探索
- 运动视差(motion parallax)这种线索是物体在视网膜成像上的运动。一般来说,对于离观察者较近的运动物体,其视网膜成像的运动量比离观察者远的物体更大,还有一个现象与运动视差有关。如果你在行驶很快的汽车里,从车窗向外看,离你很远的物体看起来似乎是跟汽车同向移动,而较近的物体似乎在向相反方向移动。
视错觉
[编辑]视错觉(Optical illusion)是指透过几何排列、视觉成像规律等手段,制作有“视觉欺骗”成分的图像引起的视觉上的错觉。视错觉一般被分为:
- 图像本身的构造导致的几何学错觉
- 由感觉器官引起的生理错觉
- 心理原因导致的认知错觉
其中特别是关于几何学的错觉,以其种类多而广为人知。
- 几何学错觉:视觉上的大小、长度、面积、方向、角度等几何构成,和实际上测得的数字有明显差别的错觉,称为几何学错觉,以下列出几个较为著名的几何学错觉:
-
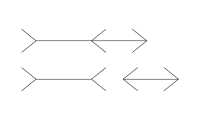 (缪莱二氏错觉):由德国社会学家弗朗茨·米勒-莱尔于1889年发现,是指两条等长平行线段,两端箭头向外的线段比两端箭头向内的线段看上去更长的现象。
(缪莱二氏错觉):由德国社会学家弗朗茨·米勒-莱尔于1889年发现,是指两条等长平行线段,两端箭头向外的线段比两端箭头向内的线段看上去更长的现象。 -
 艾宾浩斯错觉:是一种对相对大小感知的一种视错觉,以发现者,德国心理学家赫尔曼·艾宾豪斯(1850-1909)的名字命名,当两个完全相同大小的圆放置在一张图上,其中一个围绕较大的圆,另一个围绕较小的圆,而围绕大圆的圆看起来会比围绕小圆的圆要小。最近的研究表明,影响艾宾浩斯错觉感知的另外两个关键因素是周围圆圈与中心圆的距离和圆环的完整性。
艾宾浩斯错觉:是一种对相对大小感知的一种视错觉,以发现者,德国心理学家赫尔曼·艾宾豪斯(1850-1909)的名字命名,当两个完全相同大小的圆放置在一张图上,其中一个围绕较大的圆,另一个围绕较小的圆,而围绕大圆的圆看起来会比围绕小圆的圆要小。最近的研究表明,影响艾宾浩斯错觉感知的另外两个关键因素是周围圆圈与中心圆的距离和圆环的完整性。 -
 黑林错觉:于1861年由德国生理学家埃瓦尔德·黑林(Ewald Hering)发现,指两条平行直线在径向背景(如自行车辐条)前时,看起来会向外弯曲的现象,此种错觉亦称为弯曲错觉。
黑林错觉:于1861年由德国生理学家埃瓦尔德·黑林(Ewald Hering)发现,指两条平行直线在径向背景(如自行车辐条)前时,看起来会向外弯曲的现象,此种错觉亦称为弯曲错觉。 -
 加斯特罗图形:由美国心理学家约瑟夫·贾斯特罗(Joseph Jastrow)发现于1892年,两扇形虽然大小形状完全相同,但是下方的扇型看似更大。
加斯特罗图形:由美国心理学家约瑟夫·贾斯特罗(Joseph Jastrow)发现于1892年,两扇形虽然大小形状完全相同,但是下方的扇型看似更大。




- 生理错觉:主要是因为人体的视觉适应现象,人的感觉器官在接受过久的刺激后会钝化,也就造成了补色及视觉暂留的生理错觉。当视网膜上的细胞受某种色光刺激后,会对该色产生疲劳,所以在视线离开该色后,该部分的细胞暂无法作用,而未受刺激的另一部分细胞开始活动,因而产生另一种视感,也就是补色的残像。此外,过视觉疲劳而产生的视觉暂留现象就是现今动画的原理。以下列出几个著名的生理错觉例子:
-
 赫曼方格错觉:在黑色背景上的白色(或浅色)网格的交叉点上,可以看到 "幽灵般"的灰色斑点,而当直接看向一个交叉点时,灰色斑点会消失。
赫曼方格错觉:在黑色背景上的白色(或浅色)网格的交叉点上,可以看到 "幽灵般"的灰色斑点,而当直接看向一个交叉点时,灰色斑点会消失。 -
 马赫带:沿著相邻的灰色阴影之间的边界,侧向抑制使较暗的区域会假性地显得更暗,而较亮的区域则假性地显得更亮。
马赫带:沿著相邻的灰色阴影之间的边界,侧向抑制使较暗的区域会假性地显得更暗,而较亮的区域则假性地显得更亮。


- 认知错觉:这主要来自于人类的知觉恒常性属于认知心理学的讨论范围,如ASCII立体图(ASCII stereogram)、奈克方块(Necker cube)、图地反转(Negative space)及鸭兔错觉(Rabbit–duck illusion)等
前庭讯息的处理
[编辑]前庭系统vestibular system是负责人体自身平衡感和空间感的感觉系统,对于人的运动和平衡能力有关键性的作用。而前庭讯息处理使眼、头、身体能够作出补偿的运动以对抗各种机械运动。它的运作主要是由从内耳的前庭感受器出来的神经纤维,进入脑干的低级中枢,与前庭核(vestibular nuclei)的神经元形成突触连结,某些前庭神经纤维则会绕过前庭核直接进入小脑(重要的运动中枢)。
前庭也常常使人晕船或是晕机,这是身体的加速度感受器引起前庭系统兴奋所导致的现象,我们称之为晕症(motion sickness)。热也能产生相同的效果,比如说在耳朵里灌温水,会引起内耳的淋巴液运动,产生机械刺激导致头晕。因药物或抗生素中毒而伤害前庭系统的病人不会有晕症。
嗅觉讯息的处理
[编辑]嗅觉黏膜上拥有数百万个嗅觉细胞,各自只表现一种嗅觉受体;同时,单一种气味分子,可以活化不只一种嗅觉受体。因此,任何一种嗅觉,都是由不同数量及组合的嗅觉细胞,受到不同程度的活化或抑制后,将讯息传入嗅觉中枢,再经过大脑解码下的产物,非单纯一对一的关系。
嗅觉细胞为少数的双极神经元(bipolar neuron,仅有一个树突和一个轴突的神经元),其树突端往下伸入鼻腔,接收吸入的气味分子;轴突端则向上穿过头骨,进入嗅球(olfactory bulb)。带有相同受体的嗅觉细胞轴突,会在嗅球中汇集成一个嗅小球(glomerulus),每个嗅小球有25至50个来自僧帽细胞(mitral cell)的树突进驻。因此,嗅小球是嗅觉讯息的第一个整合中心。僧帽细胞将讯息进行区分及放大之后,其轴突形成嗅神经束(olfactory tract),传送至嗅觉皮质作进一步的处理。
嗅觉是所有感觉系统当中,唯一不需经过脊髓或间脑的转接,就能直接投射至前脑的感觉系统;其投射也如其他的感觉系统具地域性分布的特性,在大脑嗅觉皮质上形成地图般的构造。此外,嗅觉讯息还可以送往边缘系统(limbic system);由于边缘系统是负责情绪、记忆及行为的脑区,因此也可以解释嗅觉具有引发强烈情绪及记忆的作用。
在近年对于鼻腔的嗅粘膜和嗅球的解剖研究发现,这些部位在反映个气味上有区域性的划分,也就是说一定的区域对于一定的气味特别敏感。然而,确定各气味的感受区极端困难,因为划分气味本身就是一个难题,至今仍未找到气味分子的特定化学成分。因此,研究嗅觉的讯息处理遇上了阻碍,因为我们发现分子的大小和形状这类的化学特性和嗅觉系统的讯息编码过程没有太大的关系。
味觉讯息的处理
[编辑]味觉讯息经由三条脑神经送入脑中,分别是颜面神经中的谷索神经(chorda tympani),传送舌头前三分之一的味觉感受气的讯息;舌咽神经(glossopharyngeal nerve)传送舌头后三分之一的味觉感受气的讯息;迷走神经(vagus nerve)传送软颚和咽部的味觉感受气的讯息。这些传送味觉讯息的神经有些与延髓中的孤立核(solitary nucleus)的神经元形成突触连结;有些与桥脑中的味觉中枢的神经元形成突触连结。突触神经元的轴突将味觉讯息传到视丘,再由视丘神经元传到大脑皮质的味觉区。
味觉的讯息处理较少被研究,除了是因为人们觉得味觉比较简单外,味觉对现代人的生存已无太大影响。神经学家在味觉主要进行的细胞放电纪录主要是分析咸、酸、甜、苦的讯息编码。一派学者认为,一种细胞只反应一种味觉刺激;另一派认为味道是由众多细胞不同的兴奋模式整合所决定,称为交叉纤维模式理论(crossfiber patterning theory)。 值得注意的是,过去有学者认为舌头的特定位置会感觉到不同的味道,然而实验已证明味觉和味蕾的位置并没有明显的对应,也就是舌头上的任意位置都能感觉到不同的味道。
听觉讯息的处理
[编辑]脑的听觉构造与通路
[编辑]声音的讯号在进入耳朵后,将会经过外耳(Outer ear)、中耳(Middle ear)以及内耳(Inner ear),最后传达到内耳的纤毛细胞,以下将逐一介绍耳朵的构造。
- 外耳(Outer ear)
- 外耳包含了耳廓(Auricle)与外耳道(External acoustic),主要功能是接收外来的声能,将其传至中耳的鼓膜。耳壳就是平时我们肉眼可见的部分,可以负责收集声波,它与与外耳道口是由结缔组织相连;而外耳道则是从耳廓延伸到鼓膜,由软骨及骨骼组成,外耳道也能借由共振(resonance)的作用,来增加声音强度,使我们能对于声音频率的范围更加敏锐
- 中耳(Middle ear)
- 中耳包括了鼓膜(Tympanic membrane)、锤骨(Malleus)、砧骨(Incus)还有镫骨(Stapes),主要的机能就是放大声音讯号。鼓膜是耳朵的重要构造,它是一层较硬的膜状构造,其主要负责的工作是以振动方式传递声音,并将之传给中耳的听小骨(由锤骨、砧骨与镫骨组成),鼓膜破损的话也有可能导致传导性听力损失。鼓膜的两侧分别为外耳道及锤骨,锤骨另一侧又与砧骨相接,而砧骨的另一边也与镫骨相连,最后传至卵圆窗,而它连接著前庭。而这些构造提供了两种增强音讯的机制。首先,鼓膜到卵圆窗的面积是由大到小,能有效提高单位面积压力;再来,听小骨之间能利用杠杆原理将声波放大,来防止声波因为从外耳、中耳里的空气到内耳的耳蜗内液体(淋巴液),而造成能量的流失。而听小骨中的砧骨也与一些骨骼肌相连,而这份功能能借由肌肉的反射机制来降缓振动,以保护耳朵、听力。
- 内耳(Inner ear)
- 内耳主要是由前庭系统(Vestibular system)与耳蜗(Cochlea)组成,能将机械波振动转换成电讯号,再传到大脑中的听觉皮质,而其中前庭神经负责传送平衡觉信息,耳蜗神经负责传送听觉信息。
而每一根听神经纤维再进入脑干之后会分为两大枝,一枝到耳蜗前核(ventral cochlear nuclei),另一枝到耳蜗背核(dorsal cochlear nuclei)。从这两个核出来的纤维,一枝到上橄榄核群(superior olivary complex,接受右左两耳蜗发出的纤维,为听觉系统第一级双耳输入交互作用的地方,为听觉定位机制的重要部位),其他平行的路线汇聚在下丘核(medial geniculate nucleus),到中脑的听觉中枢。
听觉系统和其他系统一样,具有上行和下行通路,且比其他感觉系统更加明显。下列传出纤维多来自橄榄核,它们在耳蜗之中分枝很多,与每一个毛细胞的基底部形成突触连接。
音调的辨别
[编辑]我们可以听到的波频为 20-20000Hz,在这个范围内我们可以辨别极小的频率差别,在 2000Hz 以内可察觉的差别约为 2Hz。音调(pitch)和频率(frequency)不同,音调指的是感觉经验,而频率为声音的物理特性。频率不是决定音调的唯一因素,而音调的改变和频率的改变也不完全一致。辨别频率有三种理论:
- 地点理论(place theory)
- 此理论认为音调知觉决定于基底膜的波动最大的地点(“地点”应包含中枢接受兴奋的神经元地点),也就是说特定的神经元反应特定的刺激频率。
- 齐发理论(volley theory)
- 此理论强调声音刺激的频率和神经放电的模式或时间有关系(例如:500Hz 的纯音应该代表每秒 500 次的神经冲动),意味著众多神经元同时反映一个声音的刺激。单个神经元的放电模式会反映刺激的频率,刺激频率的改变也会使放电模式改变,而频率和神经冲动的一致性若由数根纤维来达成会比由单根纤维达成更为精准,因此用“齐放”来形容。
- 双重理论(duplex theory)
- 结合了地点理论和齐放理论。我们观察到基底膜的波动最大的地点与频率有关,刺激频率变了,基底膜的波动最大的地点也会改变。例如,人们观察到在某些频率的声音刺激时,耳蜗内可以完成一种傅立叶分析;不同的频率表现在基底膜不同的地段上。基底膜上的表面随著演化的过程中的加长,毛细胞、神经纤维的增加,都使得辨认听觉频率更加的精确。
在脑中,各级神经元各有其最敏感的频率。我们可以用神经元放电的时间模式来作为音频讯息的编码过程,其中包括以兴奋细胞的分布来编码(又称地址编码),以及从神经到听觉皮质的各级中枢神经元放电时间模式来编码。
声音的定位
[编辑]听觉定位的原理为,两耳之间声音强度的不同以及声音到达两耳的时间不同的情况下,两耳处理系统的结果。对于低频的声音定位,最重要的线索是声音到达两耳的时间差;而对于高频的声音定位,主要是靠两耳之间的强度差。在最好的条件下,一个人判断声源方位可以达到只有一度的误差。 人类两耳在听到声音时,声源位置的察觉可能是由中枢内许多细胞的特殊的空间和时间模式所产生,也可能是更高级中枢的某种处理结果。
在分析猫的皮质神经细胞反应时,没有发现听觉空间的特征侦察器;但在猫头鹰这类靠精准听觉定位系统来猎捕动物的鸟类脑中,在相当于下丘核的部位,发现了某些细胞的排列很像空间中的球体代表。每一个细胞都具有空间的特异性,具有像小圆椎体的感受器,圆锥尖端都辐合于猫头鹰的头部,每一个细胞只对于自己圆锥感受器的声源刺激敏感。在猫头鹰的听觉顶盖部(optic tectum),听觉和视觉的感觉系统互相符合,且所有的听觉细胞对于空间方位都相当敏感,给予确认猎物位置的双重讯号。此为猫头鹰生存所需,亦为天择的产物。
听觉双重理论 Duplex Theory
[编辑]1907年,雷利勋爵(Lord Rayleigh)受到音叉产生的单音激发,在无耳廓人头模型上研究了侧向声音定位理论。雷利提出听觉跨耳线的差异,即所谓的双重理论。 由于人的耳朵位于头部的两侧,在空间中具有不同的坐标。由于声源与耳朵之间的距离不同,两耳的声音信号之间存在时间差和强度差。这些差异被称为听觉跨耳时间差(ITD)和听觉跨耳强度差(IID)。 雷利发现: 对于低于800 Hz的声音频率,头部尺寸(即双耳间距21.5cm)小于声波的半波长。因此,听觉系统可以在两耳之间精确地确定相位延迟,而不会混淆。在这个频率范围内,听觉跨耳强度(ITD)差非常低,特别是低于约200 Hz,因此仅基于强度差异从而精确评估音源输入方向几乎不可能。
但对于高于1600 Hz的频率,头部尺寸大于声波的波长。在这些频率上,仅基于耳间相位的确定输入方向是不明确的。但是,随著耳间强度差异变大,听觉系统会针对这些强度差异进行评估。此外,两耳之间的延迟仍然可通过一些相位差异和延迟的组合检测到,这些在高频率声音时更为明显;也就是说,如果有高频声音出现,则此开始的延迟可以用于确定相应声源的输入方向。
听觉障碍及其康复
[编辑]定义
[编辑]听觉障碍,系指听觉能力受到损失,其表现可能包含无法经由听觉接收声音,或能听见声音但无法辨识声音内容、意义。
在定义上,听觉障碍 (Hearing loss) 及聋 (Deafness) 有所差异,参考《身心障碍者教育法案》 (Individuals with Disabilities Education. Act, IDEA) 对二者在孩童身上的分别定义:听力障碍被定义为“听力损伤,无论是永久性的还是波动性的,都会对儿童的教育表现产生不利影响”。耳聋被定义为“严重的听力障碍,导致儿童在透过听力(无论有或没有放大)处理语言讯息时受到损害”。
听觉障碍分级
[编辑]世界卫生组织 (World Health Organization, WHO) 依据纯音听力检查得出的结果(听力阈值),将听力障碍分为五级。而通常的“听力障碍”多指两耳听力皆受损,其分级则依优耳听觉分级为主。
| 听力障碍等级 | 相应听力测量ISO值 | 听力表现 | 建议 |
|---|---|---|---|
| 0:无损伤 | ≤25dB | 没有或非常轻微的听力问题。能够听到窃窃私语。 | 无。 |
| 1:轻微损伤 | 26-40dB | 能够在 1 m 处听到并重复以正常声音说出的单字。 | 建议咨询;可能需要助听器。 |
| 2:中度损伤 | 41-60dB | 能够在 1 m 处听到并使用提高的声音重复单字。 | 通常建议使用助听器。 |
| 3:严重损伤 | 61-80dB | 对著优耳喊叫时能够听到一些话语。 | 需要助听器。如果没有助听器,应教导唇读。 |
| 4:极严重损伤 | ≥81dB | 甚至无法听到并理解大声喊叫的声音。 | 助听器可能有助于理解单字。需要额外的复健治疗。唇读和有时手语必不可少。 |
听觉障碍的类型
[编辑]- 传达聋——外耳和中耳的病变有关的失听
- 传导聋涉及了使耳蜗兴奋的机械装置。可能是因为耳垢度塞住了外耳道、中耳炎或耳骨的错位妨碍运动等等。一般来说,中耳功能障碍造成的听力障碍或降低对一切频率的敏感性,且对高频率的敏感性丧失较多。
- 感觉神经聋——因耳蜗或听神经的受损产生的听觉障碍
- 感觉神经聋可能是因为强噪音、药物、感染、疾病、颞骨外商、显管障碍、衰老等等因素,这些情况都可能对空气传导、骨头传导声波造成障碍,使完全丧失听力。
- 药物引起的听力丧失多是受药物毒性的影响。比如说链霉素(Streptomycin)、卡那霉素(Kanamycin)、庆大霉素(Gentamicin)虽然对结核病有很好的疗效,但其中的耳中毒性质(ototoxic properties)会使病人的耳蜗和前庭受到伤害。利用组织学检查发现,链霉素会破坏基底膜底部(靠近卵圆窗的部分)以及毛细胞,使病人丧失高频率听觉,甚至完全丧失听觉且不能恢复。
- 噪音引起的听力丧失主要是因为内耳机制受到破坏。若内耳组织发生变化主要是毛细胞被破坏,且外边的毛细胞比内边的毛细胞更容易受到影响;若是长时间处于高强度的噪音下,整个柯氏器都会被破坏。有时局部伤害就算没有另外的受伤也会扩散。
- 中枢聋——由于中枢的听觉通路(脑干、视丘、大脑皮质)的损伤产生的听觉障碍
- 中枢聋往往是由脑伤所引起,但这种聋较少,因为很少有脑的两侧同时受伤。
听觉障碍的类型 受损部位 受损的原因 传达聋 外耳和中耳 耳垢堵塞、中耳炎、耳骨的错位妨碍运动 感觉神经聋 耳蜗或听神经 自然老年退化、高噪音环境、病菌感染 中枢聋 中枢的听觉通路 由脑伤所引起
- 治疗耳蜗性聋的方法
- 借由用电流刺激听神经的方式(因为毛细胞遭到破坏,但听神经仍完好)来使之兴奋,而此兴奋传入脑内后可以唤起听觉。最理想的方式是利用扩音器控制电刺激,以此模拟毛细胞产生的兴奋来诱发听神经的兴奋,虽然不能被称作治疗,但能让病患听到人们的说话声。
<补充>音乐知觉
[编辑]音乐和语言一样,都在时间结构上有明显的变化,但语言有绝对或约定成俗的意义,音乐少有此特性。音乐演奏与欣赏除依赖颞叶听觉皮层外,也依赖其它脑区,例如Premotor及Supplementarymotor cortices。聆听音乐时运动皮质的活动使音乐与舞蹈在人类文明中有紧密关系;例如人接受简单键盘音乐之演奏训练后,对其播放该音乐,不仅听觉皮质兴奋,某些运动皮质亦有活动。一个音乐家演奏熟悉的曲子(但听不见)与听该曲调(不演奏),同样都会兴奋听觉皮质,与运动皮质的某些区域。
音乐知觉与产生:音乐是一个复杂的过程,涉及许多不同的时间历程,也会激起相对应的神经反应,这些反应的产生都有一定的时间。音乐经常激起情绪反应。音乐知觉也常引起运动反应或想要动的意向,尤其是节奏明快的打击音乐。
音乐知觉—和语文的关系:音乐知觉是人类相当独特的能力。和语言一样,音乐知觉也不易找到相对应的基本单位。有一些研究探讨欣赏音乐合理解语言是否会动用同一脑区。例如:Musical structure与文法知识会启动相同的区域-布若卡氏区(Broca's area)。Musical meaning与语意结构会启动相同的区域:韦尼克区(Wernicke's Area)。音乐知觉与语言知觉的神经机制有相似也有不相似的地方。
旋律记忆:(Peretz, 1996年之研究):患者Patient C.N.丧失对旋律的记忆,但对音调的知觉正常,同时对于语言辨识也正常。
Semantic dementia(Hseih et al. 2011):右侧前颞叶受损,其知识记忆受损,连带对熟悉歌曲的旋律也受损,所以旋律可能为知识记忆一种,相对的阿兹海默症患者的损害不大。
节奏:fMRI研究显示听正常节奏相较于不正常节奏会激发运动与小脑系统的活化。随著节奏打拍子相较于找出节奏会激发基底核。
有失语症家族成员音调知觉正常,但是节奏知觉受损。前者与右脑有关而后者与左脑语言有关。
先天性音乐失能(congenital amusia):一种音乐处理能力受损的神经发育障碍,影响个体分辨音乐的能力,包括音调的辨识、音乐节奏、旋律以及和谐与否的感知。这种情况并不影响智力或一般听力,也与学习障碍或听觉问题无关。患有先天性音乐失调症的人们可能无法辨认旋律的起伏,或者无法分辨不同音乐作品或音调之间的细微差别。以实例来说,他们在唱歌或演奏乐器时可能无法分辨音准。这种失调症并非由缺乏音乐教育或文化背景的差异所引起,而是大脑处理音乐信息的方式有所不同,其右侧听觉区与额下回(Inferior frontal gyrus, IFG)可能出现了问题。值得注意的是,虽然先天性音乐失调症患者在音乐音调辨识上有明显障碍,但他们处理语言中音调的能力通常不受太大影响,特别是在使用自己的母语时。揭示了人类大脑处理音乐和语言的复杂性和部分独立的机制。 可能的治疗方法: 音乐治疗:对于先天性音乐失能患者,音乐治疗可以帮助他们加强对音乐的理解和感知能力。这种治疗通常由训练有素的音乐治疗师来进行,并可以根据患者的具体情况进行个性化的治疗计划。
听觉训练:透过听觉训练,患者可以逐步提高对音乐元素(如音调、节奏、旋律等)的识别能力。这种训练可以通过听音乐练习、听力游戏和其他听觉认知任务来实现。
认知行为疗法:认知行为疗法旨在通过改变患者的思维模式和行为来改善其心理健康状态。对于先天性音乐失能患者,这种疗法可以帮助他们处理与音乐相关的情感和挑战,并学习应对这些情感的有效方法。
神经反馈训练:神经反馈训练通常使用生物反馈技术,通过监测患者的生理指标(如脑电图、心率等)来帮助他们学习调节自己的生理过程。对于先天性音乐失能患者,神经反馈训练可以帮助他们改善与音乐相关的大脑活动模式,从而提高对音乐的感知和理解能力。
这些治疗方法可以单独应用,也可以结合使用,根据患者的具体情况和需求进行个性化的治疗计划。此外,治疗过程中的积极参与和长期执行是取得良好治疗效果的重要因素。
- 音乐知觉—音色与情绪:
- 音色(Peretz & Coltheart 模型所没有处理的):不同乐器演奏同一音调其感觉不同,这来自每个乐器对每个音符的起始过程不一,以及所伴随的不同泛音(harmonics)强度不同。Samson & Zatorre(1994):右侧颞叶损伤破坏音色知觉甚于音调。
- 情绪:音乐会引发情绪反应。节奏快的较节奏慢的音乐听起来快乐,大调快乐兴奋,小调哀伤沈痛。偏离预期的音符引发惊愕,快速规则的旋律可产生恐惧。音乐引发情绪的感觉有跨文化的效果,非洲民族能够辨识西方音乐所展现的快乐、哀伤与恐惧,纵使从来没有听过。音乐和其他刺激一样,会兴奋脑中的情绪网路。好听愉快的音乐会兴奋脑中酬赏的神经回路(Blood & Zatore, 2001)。杏仁核受损病人无法辨识恐惧表情,也无法辨识音乐的恐惧(Gosselin et al., 2007)。
- 音乐知觉—和语文的关系:
- 颞叶平面(Planum temporale)是听觉区的一部份,包含韦尼克区(Wernicke area)。
- 一般人65%左边较右边大,11%则相反。不对称性在86%的婴儿可以看出,但在左手者较不明显。被认为和语言功能有关。
- 此一解剖不对称性也存在猩猩脑中。其他语言相关区域亦然。猴子的左脑对于同类所发出的声音也叫有反应。可能为语言先驱。
- 右脑的Planum temporale则与音乐有关。但是具有绝对音感者其左侧的颞叶平面依然较大。 该区左右不对称性如下:有绝对音感的音乐家>无绝对音感的音乐家>非音乐家